迄今為止本世紀已經出現了三種冠狀病毒的全球流行。2002年11月在中國佛山報道了第一例人類感染嚴重急性呼吸系統綜合癥冠狀病毒(SARS-CoV)的病例�,到2003年7月在全球27個國家中發現了8000多例SARS病例。SARS-CoV感染的主要癥狀是流感樣��,包括發燒、頭痛���、全身乏力�、發抖和腹瀉���。SARS大流行十年后�����,沙特阿拉伯一名男子被診斷出一種新的冠狀病毒,即中東呼吸綜合征冠狀病毒(MERS-CoV),由于受感染者的國際旅行��,MERS-CoV已遍布全球�。從2012年9月到2019年12月底,總共報告了2502例經實驗室確認的MERS-CoV感染病例,包括858例相關死亡。
在2019年12月,COVID-19爆發���,當前的研究表明,這種冠狀病毒類似于SARS-CoV���。盡管這三種冠狀病毒被確定對人類具有高致病性��,但尚無有效的抗病毒治療方法。因此,當前的研究集中在快速開發疫苗和抗病毒藥物以預防和治療新冠狀病毒感染��。
目前最有吸引力的抗病毒藥物靶標之一是SARS-CoV編碼的半胱氨酸蛋白酶-木瓜蛋白酶樣蛋白酶(PLpro)�����,PLpro參與抑制細胞因子的產生和趨化因子���,負責抗病毒感染�,所以該酶是SARS-CoV抗病毒藥物設計中的重要分子靶標��。對CoV-2 PLpro底物特異性�����、結構和機制的分子了解,將通過進行合理的藥物靶向設計和研究來極大地促進有效PLpro抑制劑的研發�����。
近日《Science》上發表了一篇最新研究�����,研究人員首先使用新穎的化學方法雜交組合底物庫對SARS-CoV-2 PLpro進行了全面的活性分析,結果揭示了控制PLpro底物特異性的分子規則�����。利用這些信息��,接下來設計了具有生化特性的有效抑制劑(VIR250和VIR251)���,與其他蛋白酶相比��,它們對SARS-CoV-2 PLpro和相關SARSCoV-1 PLpro具有高選擇性。這項研究工作揭示了控制PLpro底物特異性的分子規律���,并為開發具有潛在治療價值或藥物再利用的抑制劑提供了框架。
01
底物特異性概況
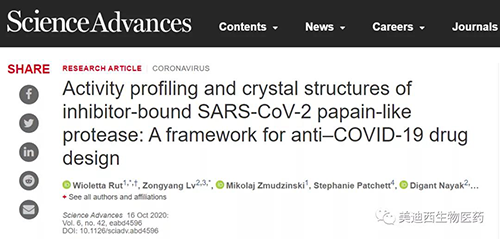
在P1位置的甘氨酸基側的肽鍵水解會導致nsp1���、nsp2和nsp3蛋白的釋放,這對于病毒復制至關重要。SARS-CoV-2 PLpro還具有去泛素化和識別這些蛋白質C末端的保守LRGG基序。先前僅針對包含天然氨基酸的組合底物庫對SARS-CoV-1 PLpro底物偏好進行的研究���,結果表明該蛋白酶識別P4-P1位置的LXGG基序����,在P3位置具有廣泛的底物特異性����。這些結果表明,對結合口袋結構更詳細的作圖應有助于新的活性底物的設計和用于抑制劑開發工作的最佳肽序列���。為了實現這個目標,研究人員開發了一個定義的組合底物庫(HyCoSuL)�����,其中包含多種非蛋白原性氨基酸���。
SARS-CoV-1和2 PLpro蛋白酶的活性分析(來源:Science Advances)
由于四肽熒光底物不能被具有去泛素化活性的酶有效地水解���,因此研究人員設計并合成了具有Ac-LRXG-ACC和HyCoSuL的一般結構的P2定義文庫�����。位置是固定的,一個位置包含19個氨基酸的等摩爾混合物��。通過設計具有針對去泛素酶的定制肽支架的文庫�����,可以在分析過程中達到每個子庫中單個熒光底物的最高可能濃度���。
P2文庫篩選顯示����,SARS-CoV和SARS-CoV-2 PLpro在此位置具有很高的底物特異性,兩種蛋白酶在P3位置均表現出廣泛的底物偏好���。SARS-CoV和SARS-CoV-2 PLpro的S3口袋不僅可以耐受帶正電的殘基,而且還可以耐受疏水性氨基酸��。這些酶無法識別酸性殘基和大多數D-氨基酸�����。SARS-CoV和SARS-CoV-2 PLpro的S4口袋只能容納疏水性殘基���,在天然氨基酸中,幾乎只能耐受亮氨酸�����。SARS-CoV PLpro在P4位置比亮氨酸更好地識別了兩個非蛋白殘基���。
02
PLpro抑制劑的開發
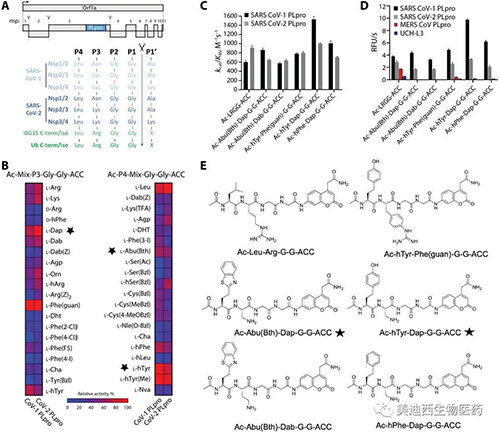
為了進一步分析具有非蛋白原性氨基酸的肽序列的選擇性��,研究人員通過交換熒光將兩種底物轉化為抑制劑標記一個反應性基團-乙烯基甲酯(VME)�。選擇VME組是因為它對DUB具有廣泛的反應性���,SARS-CoV PLpro和SARS-CoV-2 PLpro抑制劑的動力學分析結果反映了底物水解的結果��。與VIR250相比��, VIR251對這些酶的抑制作用更強,但選擇性更低��。兩種化合物均顯示出對SARS-PLpro變體的高選擇性�����,并能有效抑制SARS-CoV PLpro和SARS-CoV-2 PLpro活性����。相反�����,實際上未觀察到對人DUB UCH-L3的抑制�����,而僅觀察到對MERS-PLpro的輕微抑制。此外,將HeLa裂解物與Ub-VME一起溫育可產生交聯曲線�����,這種交聯曲線不會因滴定VIR250或VIR251而改變�����。由于已知Ub-VME的主要交聯靶標是人DUB酶,因此這些數據表明VIR250和VIR251不會與人DUB發生交叉反應,這是尋找與人DUB具有最小交叉反應性的選擇性抗病毒分子的重要發現��。
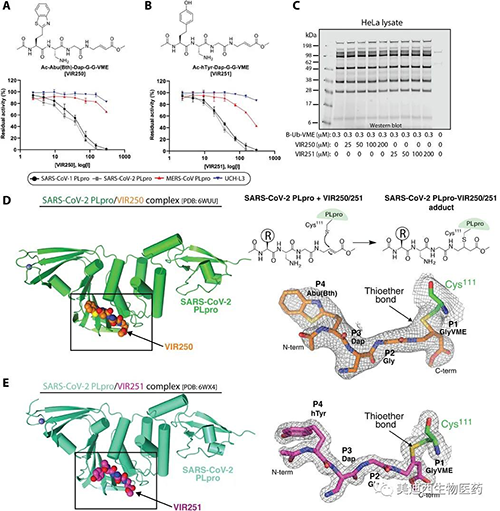
與SARS-CoV-2 PLpro配合使用的非天然氨基酸抑制劑VIR250和VIR251及其晶體結構的表征(來源:Science Advances)
接下來�����,研究人員著手確定SARS-CoV-2 PLpro與VIR250和VIR251的復合物的晶體結構���,以深入了解這些分子抑制SARS-CoV-2 PLpro活性的分子機理���,以及觀察到的底物選擇性的基礎����。大規模的交聯反應產生了CoV-2 PLpro-VIR250和CoV-2 PLpro-VIR251配合物�����,其產率和純度足以滿足衍射質量晶體的生長。
將載脂蛋白CoV-2 PLpro與CoV-2 PLpro / VIR250和CoV-2 PLpro / VIR251配合物進行比較,發現除β14-β15環位于活性位點附近并經歷構象變化外��,總體結構相似可能是由于抑制劑的結合����。結構分析表明,從CoV-2 PLpro的催化Cys 111側鏈投射出廣泛的電子密度,其中VIR250和VIR251的所有原子都可以明確地放置在其中�����。此外�����,Cys 111之間的共價鍵VIR250和VIR251都清晰可見。如預期的那樣,VIR250和VIR251抑制劑在活性位點附近均占據CoV-2 PLpro的S4-S1口袋并采用類似的結構�����,P4位置是VIR250和VIR251之間唯一的化學差異區域����,在VIR250中為Abu,在VIR251中為h Tyr����。
03
VIR250和VIR251的分子識別
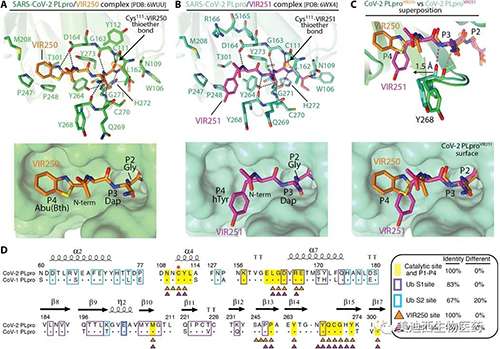
SARS-CoV-2 PLpro對VIR250和VIR251的分子識別(來源:Science Advances)
VIR250和VIR251的P3和P4位置的側鏈與CoV-2 PLpro的結合方式有重要區別����。與VIR250中的Abu(Bth)相比�,P4位置的hTyr向著S4口袋的另一側突出。β14-β15環(Asn 267�、Tyr 268和Gln 269)向VIR251的hTyr轉移1.5-?���,促進了這種新的相互作用�����,從而促進了許多新型的聯系,而這些聯系在沒有這種轉變的情況下是無法發生的����。值得注意的是,與VIR250和VIR251接觸涉及的所有CoV-2 PLpro殘基在SARS CoV-1 PLpro中都完全保守,并且兩個SARS PLpro變體的總體結構在酶的催化位點非常相似,這可能解釋了這些抑制劑具有靶向兩種酶的能力�。
就上述觀察到的底物選擇性的相關性而言��,P2對Gly的依賴性是來自CoV-2 PLpro的β14-β15和α5-α6環殘基的結果。對P3位置的正性和疏水性殘基偏愛以及對酸性殘基的選擇可能是其更寬泛的口袋和靠近Tyr 268���、Gln 269和Leu 162的酸性羰基氧的結果。在P4位置上�����,對龐大的疏水殘基的強烈偏愛可以通過P4結合口袋的疏水特性來解釋���,該特性主要由殘基Met 208����、Pro 247、Pro 248����、Tyr 264和Tyr 268形成�。值得注意的是��,SARS-CoV-2 PLpro的S4口袋的性質已被VIR250和VIR251的P4處的Abu和h Tyr側鏈所利用�。
04
CoV-1和CoV-2 PLpro處理Ub和Ubl變體
針對SARS-CoV-1 PLpro進行的研究表明�,該酶具有Ub結合域,可有效處理完整的Ub熒光底物���,因此研究人員想看看SARS-CoV-2 PLpro是否也是如此�。為此使用了基于Ub活性的探針(ABP)來標記這兩種酶����。研究人員觀察到生物素-Ub-VME對兩種蛋白酶都進行了實質性標記,其濃度是酶濃度(200 nM)的兩倍��。但是��,與SARS-CoV-2 PLpro相比,SARS-CoV-1 PLpro的標記效率更高����。接下來通過測試其與一組Ub / Ubl ABP的反應性來探索SARS-CoV-2 PLpro的Ub和Ubl蛋白底物特異性�,該分析的結果表明SARS-CoV-2 PLpro更傾向于識別ISG15和monoUb�����,并且識別K48連接的和K63連接的diUb的能力較弱��。
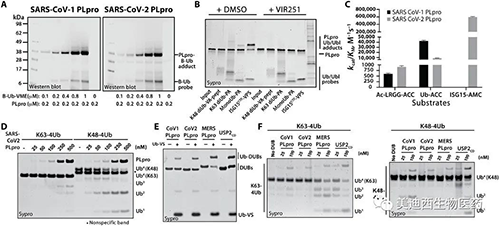
SARS-CoV-1 PLpro和SARS-CoV-2 PLpro對Ub和Ubl變體的處理(來源:Science Advances)
為了更徹底地檢查這些差異,研究人員對LRGG-ACC��、Ub-ACC和ISG15-AMC熒光底物的SARS-CoV-1和CoV-2 PLpro加工動力學進行了比較���。實驗結果表明�����,SARS-CoV-2 PLpro處理Ub-ACC的效率是SARS-CoV-1 PLpro的四倍�����,并且SARS-CoV-2 PLpro處理ISG15-AMC的效率是Ub-ACC的60倍。此外,SARS-CoV-2 PLpro與SARS-CoV-1 PLpro一樣,與K63 tetraUb相比��,對K48 tetraUb的處理更為穩健��。然而����,在并排比較中����,與SARS-CoV-1 PLpro相比���,SARS-CoV-2 PLpro表現出顯著降低了處理K48 tetraUb的能力�����。
考慮到酶之間的非常高的總體相似性����,與SARS-CoV-1 PLpro相比����,SARS-CoV-2 PLpro處理K48 polyUb鏈的能力大大降低了。為了調和這種明顯的矛盾���,研究人員將SARS-CoV-2 PLpro結構與先前報道的SARS-CoV-1 PLpro結構與K48 diUb配合使用。對SARS-CoV-1和2 PLpro催化位點的比較分析顯示�,與S1 Ub�����、VIR250和VIR251的S4-S1位置接觸時所涉及的殘基100%保守,并且與預期結構非常相似���。盡管Ub S1位點比催化位點具有更大的變異性,但總體氨基酸保守性仍然很高���,并且結構在該區域排列得很好。
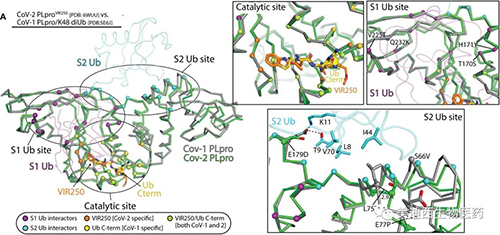
觀察到的SARS-CoV-1 PLpro和SARS-CoV-2 PLpro的Ub / Ubl加工概況的分子基礎(來源;Science Advances))
與SARS-CoV-1 PLpro的催化位點和S1 Ub位點相比�,SARS-CoV-2 PLpro的S2 Ub位點在氨基酸水平上的保守性要低得多��,并且在這些區域有一些結構差異對于分子識別S2 Ub很重要。研究人員認為SARS-CoV-2 PLpro處理K48 polyUb的能力下降的主要原因是在S2 Ub結合位點的差異,與此假設一致�,SARS-CoV-1 PLpro亮氨酸75突變為絲氨酸導致K48 diUb的結合減少了五倍����,而對monoUb沒有明顯影響���。結合SARS-CoV-2 PLpro中的其他變化���,SARS-CoV-1和2 PLpro之間在S2結合位點出現了一些相對較小的變化�,這些變化對它們處理K48 polyUb的能力的影響很大。
05
結果與討論
SARS-CoV-2 PLpro是抗病毒藥物開發的絕佳候選者��,因為它不僅阻斷病毒復制�,而且還抑制感染細胞中信號級聯的失調。通過對藥物靶向的合理設計和研究����,對SARS-CoV-2 PLpro底物特異性�、結構和機制的詳細理解將極大地促進有效PLpro抑制劑的開發��,這是此項研究的重點��。
研究人員利用獲得的有關控制SARS-CoV-2 PLpro底物選擇性的分子規則的信息來開發共價抑制劑VIR250和VIR251。這些抑制劑被證明具有活性并選擇性抑制SARS-CoV-1和-2 PLpro�,但對MERS-PLpro的活性卻弱得多���,而對人DUB UCH-L3的活性則幾乎沒有��。對于進行針對該酶的肽類抗病毒化合物的研究,這是有價值的信息�。VIR250和VIR251與SARS-CoV-2 PLpro配合使用的晶體結構揭示了它們的抑制機制�����,并為觀察到的底物特異性譜提供了結構基礎。
此外�,出乎意料的發現是�����,VIR250和VIR251的P4氨基酸占據了SARS-CoV-2 PLpro寬S4口袋的相對側,并且該口袋中還有其他區域未被任一抑制劑所結合��,這意味著未來的藥物開發工作旨在產生更多有效的抑制劑��。對SARS-CoV-2-Mpro和SARS-CoV-2 PLpro底物特異性的比較分析表明,它們具有明顯不同的底物特異性�,這表明對于肽類抑制劑����,不可能設計出一種同時作用于兩種酶的抑制劑�。但是,如果分別為這兩種蛋白酶找到了肽類抑制劑���,那么有可能需要將它們用作混合物。另一種可能的方法是尋找可以同時抑制Mpro和PLpro的小分子抑制劑�����,這種抑制劑在COVID-19的治療中肯定會非常有益�,但它可能與人半胱氨酸蛋白酶發生交叉反應,從而可能導致不良副作用。
結構分析顯示��,SARS-CoV-2 PLpro的S2 Ub結合位點存在細微的結構和序列變異����,研究人員共同認為這會削弱K48 polyUb的S2 Ub結合及隨后被加工的能力。此外,對Ub-ACC底物的酶動力學分析表明���,該酶可以有效地對其進行處理,但是在SARS-CoV-1 PLpro的情況下�,四肽底物和Ub之間的差異僅為10倍左右�。這表明兩種酶之間在外位結合區相互作用的方面與氨基酸的同一性和相似性有些不同���。鑒于Ub和ISG15偶聯在逃避宿主先天免疫反應中的作用�����,SARS-CoV-1和-2 PLpro功能的這些有趣差異是否會對病毒生物學產生任何影響尚待觀察��,并且將是未來研究的主題。
總的來說�,此項研究工作揭示了控制PLpro底物特異性的分子規則,并揭示了底物結合口袋中SARS-CoV-1和-2 PLpro之間非常高的序列和結構相似性。這些發現表明,先前發現的有關SARS-CoV-1 PLpro的信息可立即用于尋找有效的抗病毒分子,并重新靶向抑制SARS-CoV-2 PLpro的已知藥物����。此外��,與SARS-CoV-2 PLpro結合的新型抑制劑VIR250和VIR251的結構為合理開發具有增強效價和ABP的抑制劑提供了框架�����。此項研究的譜圖和晶體學研究為開發性能改善的抑制劑開辟了新途徑,研究數據也為設計一種可以同時對抗SARS-CoV-1 PLpro和SARS-CoV-2 PLpro的全選擇性抑制劑提供了幫助。美迪西作為新藥研發CRO會持續關注此項研究進展。








 相關新聞
相關新聞










